Virulence is the degree of pathogenicity within a group or species of parasites as indicated by case fatality rates and/or the ability of the organism to invade the tissues of the host. The pathogenicity of an organism - its ability to cause disease - is determined by its virulence factors. In an ecological context, virulence can be defined as the host's parasite-induced loss of fitness. Virulence can be understood in terms of proximate causes—those specific traits of the pathogen that help make the host ill—and ultimate causes—the evolutionary pressures that lead to virulent traits occurring in a pathogen strain.
The ability of a microorganism to cause disease is described in terms of the number of infecting bacteria, the route of entry into the body, the effects of host defense mechanisms, and intrinsic characteristics of the microorganism called virulence factors. Host-mediated pathogenesis is often important because the host can respond aggressively to infection with the result that host defense mechanisms do damage to host tissues while the infection is being countered.
According to evolutionary medicine, optimal virulence increases with horizontal transmission (between non-relatives) and decreases with vertical transmission (from parent to child). This is because the fitness of the host is bound to the fitness in vertical transmission but is not so bound in horizontal transmission.The pathogen population can evolve once it is in the host . There are three main hypotheses about why a pathogen evolves as it does. These three models help to explain the life history strategies of parasites, including reproduction, migration within the host, virulence, etc.
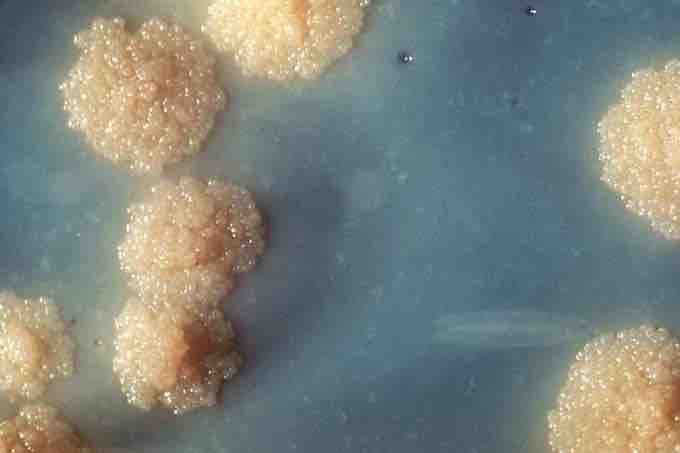
Tuberculosis Culture
The bacteria Mycobacterium tuberculosis can evolve to subvert the protection offered by immune defenses. This close-up reveals this organism's colonial morphology. Note the colorless rough surface, which are typical morphologic characteristics seen in Mycobacterium tuberculosis colonial growth. Macroscopic examination of colonial growth patterns is still one of the ways microorganisms are often identified.
- Trade-off hypothesis argues that pathogens tend to evolve toward ever decreasing virulence because the death of the host (or even serious disability) is ultimately harmful to the pathogen living inside. For example, if the host dies, the pathogen population inside may die out entirely. Therefore, it was believed that less virulent pathogens that allowed the host to move around and interact with other hosts should have greater success reproducing and dispersing. But this is not necessarily the case. Pathogen strains that kill the host can increase in frequency as long as the pathogen can transmit itself to a new host, whether before or after the host dies. The evolution of virulence in pathogens is a balance between the costs and benefits of virulence to the pathogen.
- Short-sighted evolution hypothesis suggests that the traits that increase reproduction rate and transmission to a new host will rise to high frequency within the pathogen population. These traits include the ability to reproduce sooner, reproduce faster, reproduce in higher numbers, live longer, survive against antibodies, or survive in parts of the body the pathogen does not normally infiltrate. These traits typically arise due to mutations, which occur more frequently in pathogen populations than in host populations, due to the pathogens' rapid generation time and immense numbers. After only a few generations, the mutations that enhance rapid reproduction or dispersal will increase in frequency. The same mutations that enhance the reproduction and dispersal of the pathogen also enhance its virulence in the host, causing much harm (disease and death). If the pathogen's virulence kills the host and interferes with its own transmission to a new host, virulence will be selected against. But as long as transmission continues despite the virulence, virulent pathogens will have the advantage.
- Coincidental evolution hypothesis argues that some forms of pathogenic virulence did not co-evolve with the host. For example, tetanus is caused by the soil bacterium Clostridium tetani. After C. tetani bacteria enter a human wound, the bacteria may grow and divide rapidly, even though the human body is not their normal habitat. While dividing, C. tetani produce a neurotoxin that is lethal to humans. But it is selection in the bacterium's normal life cycle in the soil that leads it to produce this toxin, not any evolution with a human host. The bacterium finds itself inside a human instead of in the soil by mere happenstance. We can say that the neurotoxin is not directed at the human host.