Genetic Alteration
In molecular biology, transformation is genetic alteration of a cell resulting from the direct uptake, incorporation and expression of exogenous genetic material (exogenous DNA) from its surroundings and taken up through the cell membrane(s) .
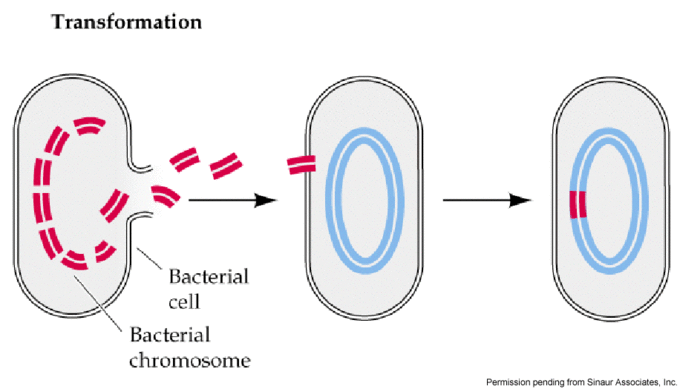
Transformation
Illustration of bacterial transformation. DNA from dead cells gets cut into fragments and exits the cell. The free-floating DNA can then be picked up by competent cells. The exogenous DNA is incorporated into the host cell's chromosome via recombination.
NATURAL TRANSFORMATION
Transformation occurs naturally in some species of bacteria, but it can also be effected by artificial means in other cells. For transformation to happen, bacteria must be in a state of competence, which might occur as a time-limited response to environmental conditions such as starvation and cell density. Transformation is one of three processes by which exogenous genetic material may be introduced into a bacterial cell; the other two being conjugation (transfer of genetic material between two bacterial cells in direct contact), and transduction (injection of foreign DNA by a bacteriophage virus into the host bacterium).
"Transformation" may also be used to describe the insertion of new genetic material into nonbacterial cells, including animal and plant cells; however, because "transformation" has a special meaning in relation to animal cells, indicating progression to a cancerous state, the term should be avoided for animal cells when describing introduction of exogenous genetic material. Introduction of foreign DNA into eukaryotic cells is often called "transfection".
Bacterial transformation may be referred to as a stable genetic change, brought about by the uptake of naked DNA (DNA without associated cells or proteins). Competence refers to the state of being able to take up exogenous DNA from the environment. There are two forms of competence: natural and artificial.
About 1% of bacterial species are capable of naturally taking up DNA under laboratory conditions; more may be able to take it up in their natural environments. DNA material can be transferred between different strains of bacteria in a process that is called horizontal gene transfer.
Some species, upon cell death, release their DNA to be taken up by other cells; however, transformation works best with DNA from closely-related species. These naturally-competent bacteria carry sets of genes that provide the protein machinery to bring DNA across the cell membrane(s). The transport of the exogeneous DNA into the cells may require proteins that are involved in the assembly of type IV pili and type II secretion system, as well as DNA translocase complex at the cytoplasmic membrane.
GRAM-POSITIVE AND GRAM-NEGATIVE DIFFERENCES
Due to the differences in structure of the cell envelope between Gram-positive and Gram-negative bacteria, there are some differences in the mechanisms of DNA uptake in these cells. However, most of them share common features that involve related proteins. The DNA first binds to the surface of the competent cells on a DNA receptor, and passes through the cytoplasmic membrane via DNA translocase. Only single-stranded DNA may pass through, one strand is therefore degraded by nucleases in the process, and the translocated single-stranded DNA may then be integrated into the bacterial chromosomes by a RecA-dependent process.
In Gram-negative cells, due to the presence of an extra membrane, the DNA requires the presence of a channel formed by secretins on the outer membrane. Pilin may be required for competence however, its role is uncertain. The uptake of DNA is generally non-sequence specific, although in some species the presence of specific DNA uptake sequences may facilitate efficient DNA uptake.
ARTIFICIAL TRANSFER
Artificial competence can be induced in laboratory procedures that involve making the cell passively permeable to DNA, by exposing it to conditions that do not normally occur in nature. Typically, the cells are incubated in a solution containing divalent cations; most commonly, calcium chloride solution under cold condition, which is then exposed to a pulse of heat shock. However, the mechanism of the uptake of DNA via chemically-induced competence in this calcium chloride transformation method is unclear.
The surface of bacteria such as E. coli is negatively-charged due to phospholipids and lipopolysaccharides on its cell surface, and the DNA is also negatively-charged. One function of the divalent cation therefore, would be to shield the charges by coordinating the phosphate groups and other negative charges, thereby allowing a DNA molecule to adhere to the cell surface. It is suggested that exposing the cells to divalent cations in cold condition may also change or weaken the cell surface structure of the cells making it more permeable to DNA. The heat-pulse is thought to create a thermal imbalance on either side of the cell membrane, which forces the DNA to enter the cells through either cell pores or the damaged cell wall.
Electroporation is another method of promoting competence. Using this method, the cells are briefly shocked with an electric field of 10-20 kV/cm which is thought to create holes in the cell membrane through which the plasmid DNA may enter. After the electric shock, the holes are rapidly closed by the cell's membrane-repair mechanisms.
O. T. Avery, et al. were first to demonstrate that "rough" colonies of S. pneumoniae could be transformed to "smooth" (capsule producing) colonies by addition of DNA extracts of the former to the latter, thus "transforming" them. (See Lederberg below)
- Lederberg, Joshua (1994). The Transformation of Genetics by DNA: An Anniversary Celebration of AVERY, MACLEOD and MCCARTY(1944) in Anecdotal, Historical and Critical Commentaries on Genetics. The Rockfeller University, New York, New York 10021-6399. PMID 8150273.