The Double-Stranded DNA (dsDNA) tailed phages, or Caudovirales, account for 95% of all the phages reported in the scientific literature, and possibly make up the majority of phages on the planet. Nineteen families that infect bacteria and archaea are currently recognized; of these, 15 have double-stranded DNA genomes.
Under the Baltimore classification scheme, the Caudovirales are group I viruses as they have double-stranded DNA (dsDNA) genomes, which can be anywhere from 18,000 base pairs to 500,000 base pairs in length. The virus particles have a distinct shape; each virion has an icosohedral head that contains the viral genome, and is attached to a flexible tail by a connector protein. The order encompasses a wide range of viruses, many of which contain genes of similar nucleotide sequence and function. Some tailed bacteriophage genomes can vary quite significantly in nucleotide sequence, however, even among the same genus. Due to their characteristic structure and possession of potentially homologous genes, it is believed these bacteriophages possess a common origin. There are at least 350 recognized species in this order.
Upon encountering a host bacterium, the tail section of the virion binds to receptors on the cell surface and delivers the DNA into the cell by use of an injectisome-like mechanism (an injectisome is a nanomachine that evolved for the delivery of proteins by type III secretion). The tail section of the virus punches a hole through the bacterial cell wall and plasma membrane and the genome passes down the tail into the cell. Once inside, the genes are expressed from transcripts made by the host machinery, using host ribosomes. Typically, the genome is replicated by use of concatemers, in which overlapping segments of DNA are made, and then put together to form the whole genome.
Viral capsid proteins come together to form a precursor prohead, into which the genome enters. Once this has occurred, the prohead undergoes maturation by cleavage of capsid subunits to form an icosohedral phage head with 5-fold symmetry. After the head maturation, the tail is joined in one of two ways: either the tail is constructed separately and joined with the connector, or the tail is constructed directly onto the phage head. The tails consist of helix-based proteins with 6-fold symmetry. After maturation of virus particles, the cell is lysed by lysins, holins, or a combination of the two.
Because the lack of homology between the amino acid and DNA sequences of these viruses precludes these from being used as taxonomic markers (as is common for other organisms), the three families here are defined on the basis of morphology. This classification scheme was originated by Bradley in 1969 and has since been extended. All viruses in this order have icosahedral or oblate heads, but differ in the length and contractile abilities of their tails. The Myoviridae have long tails that are contractile, the Podoviridae have short noncontractile tails, and the Siphoviridae have long non-contractile tails. Siphoviridae constitute the majority of the known tailed viruses .
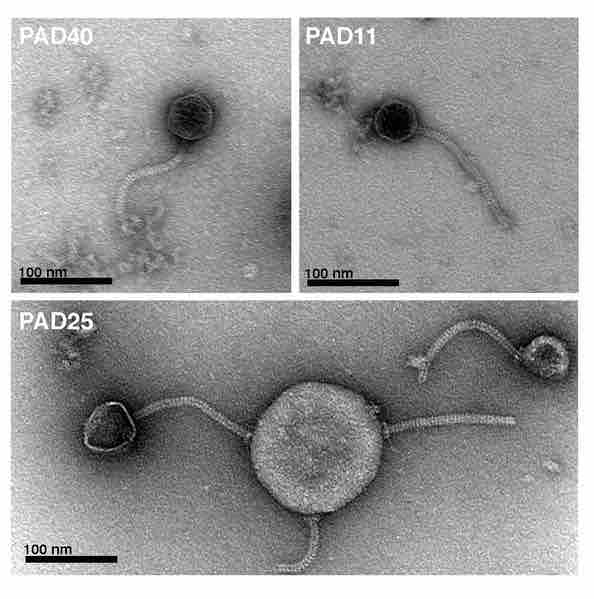
Siphovirus phages
Electron micrographs of bacteriophages from P. acnes. Phages were negatively stained with 0.75% uranyl formate and subjected to transmission electron microscopy. The phages have a head of approximately 55 nm in diameter, loaded with genetic material. Their tails have a size of 150 × 10 nm and are flexible and non-contractile. In the lower micrograph, PAD25 is adhering to bacterial cell debris, and two phages have lost their heads. At the attachment site between the phage and the cell debris, a base plate with attached spikes can be observed. All phages were classified as Siphoviruses based on their morphology.